

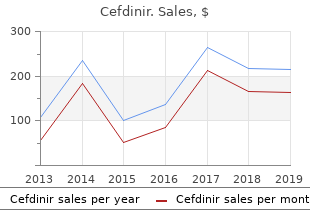
University of Michigan-Dearborn. I. Ines, MD: "Purchase cheap Cefdinir - Quality online Cefdinir no RX".
A recent survey of the literature found nonrandom associations between parasite geno- typeswithinhosts (Lord et al order 300 mg cefdinir free shipping antibiotics starting with z. For sexual parasites cheap 300 mg cefdinir with visa antibiotic with sulfa, nonrandom associations within hosts often affect mating patterns effective cefdinir 300 mg bacteria jacuzzi. Mating typically occurs between the parasites within a host or between parasites in a vector that were recently derived from one or a few hosts. Nonrandom mating alters heterozygosity at individual loci and the opportunities for recombination between loci. Host immunity may influence the distribution of strains within hosts. Theirdata suggest that the two variants mutually interfere with T cell attack against the parasite, so both variants do betterinthehost when they are together. In general, the immunological profile of each host constrains the range of parasite variants that may coinfect that host. EFFECTIVE POPULATION SIZE The number of adult genotypes sampled to produce the progeny gen- eration influences the effective size of the population (Wright 1969; Crow and Kimura 1970; Li 1976; Hartl and Clark 1997; Hedrick 2000). Population size affects many statistical properties of genetic structure. For example, suppose a particular parasite genotype sweeps through ahost population, causing a widespread epidemic. This epidemic ge- notype rises to a high frequency as other genotypes fail to spread or decline in abundance. Descendants of the population after an epidemic will likely come from theepidemic genotype (Maynard Smith et al. The effec- tive size of the population is small because of the limited number of ancestral genotypes. The spread of an epidemic genotype carries along in strong association the alleles of that genotype atdifferent loci. Conse- quently, strong genome-wide linkage disequilibrium may appear when descendants of the epidemic genotypes are sampled among genotypes descended from other lineages (Maynard Smith et al. Population size also influences the pattern of genomic evolution by natural selection (Kimura 1983). When the effective population size is small, chance events of sampling canfavorone allele over another. This stochastic sampling reduces the power of natural selection to shape evo- lutionary patterns of antigenic variation. Statistical descriptions of the sampled data readily allow calculation of heterozygosity levels at single loci, the linkage disequilibrium between loci within genomes, and the spatial distribution of genotypes. Yes, but only if we can rule out alternative processes that could lead to the same pattern. Suppose, for example, that we demonstrate genome-wide linkage dis- equilibrium. The pattern by itself is interesting, because we have es- tablished that the parasites fall into discrete strains. Each strain can be identified by its combination of alleles, allowing the movement of strains to be followed. Each strain can also be studied for its unique antigenic and physiological properties, such as response to drugs. The pattern of genome-wide linkage does not tell us what process cre- ated that pattern. The pattern may be created by frequent epidemics, each epidemic stemming from a limited number of genotypes. The par- asite may be asexual, binding together alleles at different loci because no process mixes allelesbetween genotypes. Or, sex and the physical mixing of genotypes by recombination may occur in every generation, but with all mating confined to the pool of genotypes within each host. If only one parasite genotype typically infects a host, then all mating occurs between members of the same lineage with no opportunity for recombination to break down associations between loci. One can carefully list all processes that could lead to the observed pattern and then do statistical tests of the data to distinguish between the potential causes. Most statistical analyses have not focused on antigenic variation. In- stead, those analyses have used dataongenetic variability from loci sampled across the genome. In some cases, the analyses use common enzyme (housekeeping) loci (Enright and Spratt 1999). Housekeeping loci are likely to evolve relatively slowly compared with other parts of the genome. The relatively slow rates of change provide a good indi- cator of common ancestrybetween genomes that have been separated for long periods of time. Other analyses use rapidly evolving loci,which provide more information about recent divergence from common ances- tors (Tibayrenc 1999). Ireview some population studies of genetic structure. I emphasize only the background needed for understanding antigenic variation, leav- ing out much of the analytical detail. I start with linkage of alleles across the entire genome. Four different barriers prevent genetic mixing (Maynard Smith et al. First, asexual reproduction separates lineages irrespective of geograph- ical or ecological locality. Differentiated strains will occur jointly in the same area. In addition, particular multilocus combinations of genes may disperse widely and be found in different regions without being broken up by recombination with local varieties. Second, physical separation by geography or habitat prevents genetic mixing. Geographic subdivision is common in many populations. Eco- logical subdivision may arise if some genotypes occur mainly in one host species, whereas other genotypes are confined to a different host. Sex- ual species divided by physical barriers will have mixed genomes within local regions and differentiated genomes across barriers. Particular mul- tilocus genotypes are unlikely to be found far from their native region because they will be broken up by recombination with neighboring ge- notypes. Third, demography can separate lineages if each host or vector car- ries only a single parasite genotype.
Diseases
Patients with HBV reactivation were more likely to Yasuhito Tanaka (Department of Virology and Liver Unit cefdinir 300mg line infection viral, Nagoya have a poorer prognosis order 300 mg cefdinir with mastercard bacteria kingdom classification. Ryuzo Ueda (Department of Tumor Immunology discount cefdinir 300 mg with amex antibiotics for uti gonorrhea, Aichi tive HBV clones such as precore mutants. Chugai Pharmaceutical Company for providing clinical data on 211 These 2 studies also showed that most HBV reactivation was patients developing serious hepatitis B after rituximab-containing observed within 1 year after completion of rituximab-containing chemotherapy. Frequency of hepatitis B virus Conflict-of-interest disclosures: K. Mortality secondary to Kensei Tobinai, MD, PhD, Department of Hematology, National fulminant hepatic failure in patients with prior resolution of hepatitis B Cancer Center Hospital, 5-1-1 Tsukiji, Chuo-ku, Tokyo 104-0045, virus infection in Japan. Phone: 81-3-3542-2511; Fax: 81-3-3542-3815; e-mail: 20. Lok AS, Liang RH, Chiu EK, Wong KL, Chan TK, Todd D. Reactivation of hepatitis B virus replication in patients receiving 21. Reverse seroconversion of preemptive lamivudine therapy for hepatitis B patients undergoing hepatitis B after allogeneic bone marrow transplantation: a retrospective chemotherapy. Hepatitis B virus reactivation in B-cell N Engl J Med. Hepatitis B virus reactivation in immunosuppressive therapy: evaluation of both HBsAg-positive and lymphoma patients with prior resolved hepatitis B undergoing antican- HBsAg-negative cohorts. B virus following systemic chemotherapy for malignant lymphoma. Rituximab-associated hepatitis hepatitis B virus infection. B virus (HBV) reactivation in lymphoproliferative diseases: meta- 28. Prevention of hepatitis B virus analysis and examination of FDA safety reports. FDA: Increased HBV reactivation risk with ofatumumab or 29. Management of patients with hepatitis B who rituximab. EASL clinical practice targeted therapy with rituximab in patients with rheumatoid arthritis. A revisit of prophylactic lamivudine for granulomatosis with polyangiitis (Wegener’s granulomatosis): compari- chemotherapy-associated hepatitis B reactivation in non-Hodgkin’s son of efficacy in granulomatous versus vasculitic manifestations. Clinical and virological mide for ANCA-associated vasculitis. A comparison of entecavir and CD20 immunotherapy in fludarabine-refractory chronic lymphocytic lamivudine for HBeAg-positive chronic hepatitis B. Tenofovir disoproxil fumarate patients with CLL and coexisting conditions. Efficacy of entecavir treatment virus persists for decades after patients’ recovery from acute viral for lamivudine-resistant hepatitis B over 3 years: histological improve- hepatitis despite active maintenance of a cytotoxic T-lymphocyte ment or entecavir resistance? Retrospective and prospective therapy: results of interim analysis [abstract]. Blood (ASH Annual studies of hepatitis B virus reactivation in malignant lymphoma with Meeting Abstracts). Chemotherapy-induced hepatitis B entecavir prophylaxis for rituximab-associated hepatitis B virus reactiva- reactivation in lymphoma patients with resolved HBV infection: a tion in patients with lymphoma and resolved hepatitis B. Is Antiviral Prophylaxis observational study of hepatitis B virus (HBV) DNA monitoring and Necessary to Prevent Hepatitis B Virus (HBV) Reactivation in Patients preemptive antiviral therapy for HBV reactivation in patients with With HBV-Resolved Infection Receiving Rituximab-Containing Chemo- B-cell non-Hodgkin lymphoma following rituximab containing chemo- therapy? Grupp1,3 1Division of Oncology and 3Department of Pathology, The Children’s Hospital of Philadelphia, and Department of Pediatrics, University of Pennsylvania Perelman School of Medicine, Philadelphia, PA; and 2University of Texas M. Anderson Cancer Center, Houston, TX Relapsed and refractory leukemias pose substantial challenges in both children and adults, with very little progress being made in more than a decade. Targeted immunotherapy using chimeric antigen receptor (CAR)-modified T cells has emerged as a potent therapy with an innovative mechanism. Dramatic clinical responses with complete remission rates as high as 90% have been reported using CAR-modified T cells directed against the B-cell-specific antigen CD19 in patients with relapsed/refractory acute lymphoblastic leukemia. Supraphysiologic T-cell proliferation, a hallmark of this therapy, contributes to both efficacy and the most notable toxicity, cytokine release syndrome, posing a unique challenge for toxicity management. Further studies are necessary to identify additional targets, standardize approaches to cytokine release syndrome management, and determine the durability of remissions. Learning Objective Design and mechanism CARs were first described 20 years ago as a means of introducing ● To describe factors leading to successful and highly active 12 tumor specificity into adoptive cell therapy. The principle of cell therapies and the risk factors for toxicity antigen-specific T cell therapy was realized with first-generation CARs, which link an antibody-derived single-chain variable frag- ment (scFv) to the CD3 intracellular signaling domain of the TCR Introduction complex (Figure 1). Subsequent modifications incorporated 1 Relapsed acute lymphoblastic leukemia (ALL) is a leading cause of (second-generation CAR) or 2 (third-generation CAR) costimula- cancer deaths in children and has a dismal prognosis in adults. Retroviral and lentiviral vectors allow (CR2); however, those remissions are frequently not sustained. Salvage therapy for second or greater BM relapse induces remis- The advantage of this approach is the potential for long-term disease sions in only 40% of patients and long-term survival is quite poor. However, Overall survival for adults with ALL is poor (30%–40%),5 and ongoing on-target toxicity and the theoretical risk of transformation induction of CR2 remains quite difficult in adults, with rates of 50% are potential concerns. In recent years, genomic characterization has integration into the genome is a concern. RNA-based approaches guided the study of therapies targeting leukemogenic lesions,6,7 can produce substantial tumor responses; however, expression which were driven by the success of imatinib in Philadelphia beyond 1 week requires repeated infusions,19 so long-term disease chromosome (Ph)-positive ALL8; however, driver lesions can be control may still be possible with this approach but would require found in only a subset of ALL. Immune-mediated elimination of tumor cells has long been recog- Regardless of the method of gene transfer, in vitro cell culture systems nized and is the basis for both cancer vaccines and cellular therapies, for T cell expansion are used to manufacture large quantities of including hematopoietic stem cell transplantation (HSCT). These systems use antibodies and/or tive transfer of T cells engineered to express a chimeric antigen various artificial APCs to engage CD3 and activate T cells, with receptor (CAR) is emerging as an extremely powerful technology costimulation provided by a second signal or cytokine. This chapter differentiated and have minimal replicative capacity. CAR molecules link an extracellular scFv to intracellular signaling domains. The intracellular component includes the CD3 intracellular signaling domain of the TCR either alone (first-generation) or in combination with 1 (second-generation) or 2 (third-generation) costimulatory domains. Once engaged, Leukemia targets CARs link activated T cells to malignant cells expressing the target The ideal target for CAR-modified T cells would be universally antigen, triggering a cell-mediated immune response that bypasses expressed on tumor cells but not expressed on normal cells. Engagement can lead to a cytotoxic T-cell response as such targets are rare, antigens that are minimally expressed on well as massive T-cell proliferation in vivo. Peripheral blood mononuclear cells collected by leukapheresis are expanded ex vivo and transduced to express the CAR molecule before infusion into the patient. In this example, magnetic beads coated with antibodies to CD3 and CD28 are used for ex vivo expansion. CD19 has emerged as an attractive target due to its specificity for one cell lineage (B cells) and near universal expression on B-cell malignancies, including chronic lymphocytic leukemia (CLL), ALL, and many non- Hodgkin lymphomas. Although individual leukemias may aberrantly express anti- gens that are not normally expressed on T cells, there is no universal target that is specific to T lymphoblasts.

Diseases
However buy cefdinir cheap online virus 68 symptoms, antibodies that bind too strongly clear the matching antigens quickly and prevent feedback stimulation to their B cells cefdinir 300mg fast delivery antimicrobial medicines. The later phases of B cell competition and maturation of IgG favor antibodies with increased on-rates of associa- tion to epitopes rather than increased equilibrium binding affinity cheap cefdinir online visa virus zapadnog nila simptomi. The second section discusses cytotoxic T lymphocyte (CTL) immuno- dominance. Aspects of specificity such as MHC binding and avoidance of self-recognition determine which epitopes could potentially be rec- ognized. Among this potential set, some epitopes dominate others in stimulating a CTL response. Earlier stimulation of T cell lineages in response to infection rather than more rapid T cell division seems to determine the dominance of lineages. Dominant lineages may repress subdominant lineages bypushing the abundance of pathogens below the threshold needed to trigger weaker, subdominant responses. The third section describes original antigenic sin, in which the speci- ficity of the immune response depends on the sequence of exposure to antigenic variants. If a host first encounters a variant A and then alatervariant A ,thesecond variant will sometimes restimulate the initial response against A rather than a new, specific response against A. Inthiscase,A recalls the memory against an earlier cross-reacting epitope rather than generating a primary, specific response against it- self. Sometimes the cross-reaction is rather weak, causing the host to 74 CHAPTER 6 respond weakly to the second antigen because of interference by its memory against the first variant. Original antigenic sin has been ob- served in both antibody and CTL responses. The final section takes up promising issues for future research. The initial antibody response, detected one week after injection into a mouse, contained heterogeneous IgM against several epitopes that collectively spanned theentire 100-amino-acid sequence. By contrast, the IgG response four weeks after injection was highly specific for a single epitope. These ob- servations support the idea that the naive antibody repertoire can bind almost any epitope, but that only a subset of the initially binding anti- bodies stimulate their B cell clones to expand significantly and make the transition to IgG production. REVIEW OF PROCESSES BY WHICH ANTIBODY RESPONSE DEVELOPS Major expansion of a B cell clone and transition to IgG production typ- ically depend on stimulation from helper T cells, although some nonpro- tein antigens can stimulate IgM response without T cell help (Janeway et al. The interaction between B cells and T cells happens roughly as follows. The B cell receptor (BCR) is an attached form of antibody, which has specificity for particular epitopes. Each B cell expresses many BCRs on its surface, each with the same specificity. When a BCR binds antigen, it maypullthe antigen into the cell. If the antigen is a pro- tein, the B cell processes the antigen into smaller peptides, binds some of those peptides to MHC class II molecules, and presents the peptide– class II complexes on the cell surface. If a helper (CD4+)TcellhasaTcellreceptor (TCR) that binds the peptide–class II complex, then the T cell sends a stimulatory signal to IMMUNODOMINANCE WITHIN HOSTS 75 the B cell. Thus, B cell stimulation requires binding to an epitope of an antigen, processing the antigen, and finding a helper T cell that can bind an epitope of the same antigen. The epitopes recognized by the BCR and TCR may differ, but must be linked on the same antigen molecule to providematches to both the BCR and TCR (Shirai et al. T cell stimulation causes B cells to divide more rapidly, to undergo somatic hypermutation, and to switch from IgM to IgG production. Immuno- dominance arises when some B cells receive relatively greater stimula- tion from helper T cells. Signal strength depends on the dynamics of antigen binding forBCRsandTCRs. The vertebrate host has specialized organs to facilitate interaction be- tween B and T cells. The initial interaction occurs when antigen-binding Bcells are trapped in a zone of lymphoid tissue that has a high density of T cells. Some of the stimulated B cells differentiate into antibody fac- tories, whereas others migrate along with matching T cells to primary follicles of the lymphoid tissue. There, if the B cells receive sufficient stimulation from T cells, they undergo rapid division to form germinal centers. At these centers, the B cells hypermutate and proceed through affinity maturation. AFFINITY WINDOW FOR EPITOPE-PARATOPE BINDING The naive B cell repertoire binds with varying affinity to different epi- topes of an antigen. The relative stimulation of different B cell clones by an antigen determines progression to the next steps in B cell response. Stimulation depends on the affinity of the BCR paratopes (binding sites) for their particular epitopes. Rao (1999) found an affinity window for stimulation of B cells. Very strong epitope-paratope binding prevents stimulation; weakly binding Bcells are outcompeted for stimulatory signals. One of these epitopes stimulated the immunodominant IgG response; the other wasatthe opposite end of the peptide. I refer to the immunodominant epitope as D and the subdominant epitope as S. Surprisingly, the early antibody response was stronger against S 76 CHAPTER 6 than D. However, secreted antibodies against S bound so efficiently to Sthatthey outcompeted the matching BCR and prevented stimulation of the B cell lineage. By contrast, anti-D antibodies bound with lower affinity and did not outcompete the matching BCR, allowing that B cell lineage to receive strong stimulation from the antigen. They began by constructing a peptide that had on one side a known B cell antigen of hepatitis B virus and on the other side a known T cell epitope from the malaria parasite Plasmodium falciparum. Theyinjected this chimeric peptide into mice and followed the antibody response. The early IgM response had specificities that spanned the entire hepatitis B segment. By contrast, the later IgG response focused on a single epitope in the hepatitis B segment composed of the four amino acids DPAF. Single amino acid changes in DPAF destroyed immunodominance by this epitope, causing nearby epitopes to dominate the IgG response. The anti-DPAF antibodies had affinities between 8- and 60-fold higher than antibodies against neighboring epitopes. Immunodominance depended on competition for antigen-specifichelperTcells, which arelimiting during the initial stages of an immune response.