

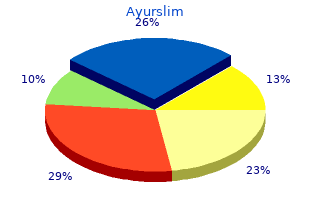
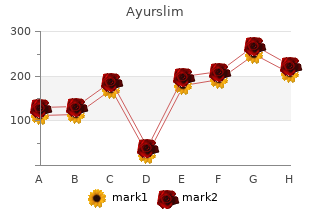
Washington Bible College / Capital Bible Seminary. X. Tippler, MD: "Purchase Ayurslim no RX - Discount Ayurslim OTC".
The superior surface of the bladder is separated by peritoneum from part of the sigmoid colon and from coils of small intestine (33 cheap ayurslim 60 caps on-line herbs pool. They are separated from these bones by a mass of fat and by the puboprostatic ligaments (see below) cheap 60 caps ayurslim fast delivery kan herbals relaxed wanderer. The base of the bladder lies in front of the rectum discount ayurslim online mastercard herbs like weed, but is partly separated from it by the right and left seminal vesicles and the right and left ductus deferens (33. Traced anteriorly this perito- neum becomes continuous with that lining the anterior abdominal wall. In the middle line this peritoneum is raised into a fold called the median umbilical fold because of the presence here of the median umbilical ligament. Traced laterally the peritoneum of the superior surface is refected on to the lateral pelvic wall. Traced posteriorly the peritoneum on the superior surface of the bladder passes on to the upper part of the base. The peritoneum lined depression between the urinary bladder and the rectum is called the rectovesical pouch (37. In the fetus the rectovesical pouch is much deeper and extends up to the pelvic foor. The lower part of the pouch is obliterated by fusion of the layers of peritoneum lining it. The remains of this peritoneum persist as the rectovesical fascia that separates the lower part of the base of the bladder, and lower down the prostate, from the rectum. Chapter 33 � Pelvic Viscera and Peritoneum 653 Relations of Urinary Bladder in the Female 1. The greater part of the superior surface of the bladder is lined by peritoneum that separates it from the body of the uterus (33. When traced backwards this peritoneum is refected on to the front of uterus at the junction of the body with the cervix. The posterior part of the superior surface of the bladder is in direct contact with the upper part of the cervix. The relations of the inferolateral surfaces of the bladder are the same as in the male except that the pubopros- tatic ligaments are replaced by the pubovesical ligaments. Ligaments of the Urinary Bladder the urinary bladder is kept in place by a number of so-called ligaments. The median umbilical ligament connects the apex of the urinary bladder to the umbilicus. It is the remnant of an embryonic structure the urachus that is derived from the allantoic diverticulum. The fascia over the upper surface of the levator ani (pelvic fascia) is thickened anteriorly to form the medial and lateral puboprostatic ligaments (in the male) or the pubovesical ligaments (in the female). Laterally the same fascia stretches from the bladder to the fascia covering the obturator internus. The lateral margins of the base of the bladder are joined to the lateral pelvic wall by fascia surrounding the veins that pass from the bladder to the internal iliac veins. The median umbilical ligament raises up a median fold of peritoneum called the median umbilical fold (33. In the fetus the right and left umbilical arteries pass from the internal iliac arteries to the umbilicus (on their way to the placenta). Their distal parts become obliterated and form the medial umbilical ligaments that connect the superior vesical arteries to the umbilicus. They raise up folds of peritoneum called the right and left medial umbilical folds. Peritoneum refected from the superior surface of the bladder to the lateral wall of the pelvis is referred to as the lateral false ligament of the bladder. Two folds of peritoneum (right and left) pass backwards from the lateral margin of the base of the bladder to the sacrum. These folds pass lateral to the rectum and form the lateral boundaries of the rectovesical pouch. These folds are called the sacrogenital folds or the posterior ligaments of the bladder (33. On the posterior wall of the bladder, however, there is a triangular area where the mucosa is relatively fxed. The ureters open into the urinary bladder at the upper lateral corners of the trigone while the upper end of the urethra opens at the lower angle. The upper margin of the trigone forms a ridge stretching between the openings of the two ureters. The urinary bladder is supplied (in the male) by the superior and inferior vesical arteries. In the female the inferior vesical artery is replaced by the vaginal artery and the uterine artery also gives branches to the bladder. Veins from the bladder pass backwards in the posterior ligaments of the bladder to reach the internal iliac veins. Parasympathetic nerves stimulate the detrusor muscles and are inhibitory to sphincters. However, it is now believed that normal bladder function is controlled only by parasympathetic nerves and that sympathetic nerves only have a vasomotor effect. Sensations of bladder flling and pain travel through both sympathetic and parasympathetic nerves. Within the central nervous system pathways for sensations of bladder flling and for pain are different. Pain from the bladder can be abolished by anterolateral cordotomy without affecting sensations of blad- der flling. Fibres travel through pelvic splanchnic nerves, inferior hypogastric plexus and vesical plexus. The overlying anterior abdominal wall is also absent so that the posterior wall of the bladder (trigone) appears on the surface of the body. The lumen of the bladder may be divided completely (by septa) or partially (by a constriction) into upper and lower compartments. In an infant the urinary bladder is partially in contact with the anterior abdominal wall. In the adult the empty bladder does not come in contact with the anterior abdominal wall. However, when it is distended its upper part is in contact with the abdominal wall above the pubic symphysis. It is important to note that as the distended bladder ascends the fold of peritoneum passing from the anterior abdominal wall to the superior surface of the bladder also rises so that no peritoneum intervenes between a distended bladder and the anterior abdominal wall. In a patient with urinary obstruction, and consequent distension of the bladder, the distension can be relieved by passing a needle into the bladder through the anterior abdominal wall (just above the pubic symphysis). The bladder can be approached surgically through a suprapubic incision (after distending it). This opera- tion is used for removal of stones from the bladder (suprapubic lithotomy).
Syndromes
The first hypothesis suggests that the membrane may be a part of endoplasmic reticulum or Golgi apparatus (Yorimitsu and Klionsky 2005) order cheap ayurslim online juvena herbals. According to the second hypothesis buy 60caps ayurslim free shipping himalaya herbals acne-n-pimple cream, the phagophore is synthesized in the cytoplasm de novo (Yang et al best buy for ayurslim lotus herbals quincenourish review. Most researchers working on the autophagy support the former hypothesis because the transmembrane protein Atg9 is localized in membranes of the late endosomes and in the trans vesicles of the Golgi apparatus. During starvation, Atg9 circulates between Golgi apparatus or late endosomes and newly formatted isolation membrane, providing compounds necessary for autophagosome creation (Yang et al. The Atg9 protein is present in the phagophore, but it could not be detected on the surface of already formed autophagosomes (Kost et al. This is caused by removal of this protein from the membrane by the Atg1 protein, which is a serine-threonine kinase, a component of the Atg1-Atg13-Atg17 complex. Nucleation and elongation At the early stage of the isolating membrane formation, i. On the other hand, proteins from the Bcl-2 family (B-cell leukemia/lymphoma-2) and the protein Ambra1 (activating molecule in Beclin1-regulated autophagy) are negative regulators. In the next step of the isolating membrane elongation, there are two conjugation processes which require the involvement of two protein complexes. The first one is Atg5-Atg12- Atg16L, which requires Atg7 and Atg10 for its formation. Then, it is transferred to Atg10, also forming a thioester bond, and finally it is conjugated with Atg5 through an amide bond. This bond appears to be un-reversible, as no protease able to cut the Atg12-Atg5 complex could be found (Yang et al. At the later step, the Atg16L protein is attached to the Atg12-Atg5 protein, forming a covalent bond with Atg5. The Atg12-Atg5-Atg16L complex oligomerizes, forming structures that are used during elongation of the isolating membrane, which results in appearance of the pre-autophagosomal membrane (Yang and Klionsky 2010). Initially, the proteins are attached uniformly on the membrane when it is being formed, however, as the phagophore is being elongated, they move towards its outer surface and then they dissociate when the autophagosome is complete. Activation of the Atg8 protein is initiated due to removal of the C-terminal amino acid, alanine, by the cysteine protease Atg4. The Atg8 protein is transferred on the Atg3 protein, forming another thioester bond. After the fusion of autophagosome with lysosome, the amide bond is hydrolyzed by the Atg4 protease, and Atg8 dissociates into the cytoplasm (Marino and Lopez-Otin 2004). It was initially considered as a microtubule-associated protein (microtubule associated protein 1 light chain 3). These processes cause formation of the autophagosome from the isolating membrane (phagophore), the structure containing a fragment of the cytoplasm together with some proteins and organelles. Fusion of autophagosome with lysosome After formation of the autophagosome, the outer membrane of autophagosome and lysosomal membrane fuse and form autophagolysosome (sometimes called autolysosome). Autophagosome fuses first with early and late endosomes which provide not only compounds to be degraded, but also factors required for the fusion between autophagosome and lysosome. It appears that endosomes lower the pH inside autophagosomes, creating favorable conditions for actions of lysosomal hydrolases (Glick et al. Mutations in the gene coding for the Rab7 protein impair the fusion of autophagosomes with late endosomes and lysosomes (Gutierrez et al. It was demonstrated that compounds which destabilize microtubules also inhibit autophagosome maturation. In cells treated with cytochalazine D, which blocks actin polymerization, a decrease in number of autophagosomes and autophagolysosmes was observed. Moreover, after treatment with nocodazole, which interferes with microtubule dynamics, the fusion of autophagosome with lysosome was blocked (Kochl et al. On the other hand, taxol, which stabilizes microtubules, caused an increase in efficiency of autophagolysosome formation (Marino and Lopez-Otin 2004; Yang et al. Lysosomal degradation Inside the autophagolysosme, there is an acid environment, which assures optimal pH for action of acid hydrolases which digest compounds enclosed inside this structure, together with its internal membrane. Thus, autophagolysosomes become single-membrane structures containing degraded compounds of the cytoplasm (Roy and Debnath 2010). Go to: Autophagy activation pathways In the regulation of the autophagy process, there are several signaling pathways. It plays roles not only in autophagy regulation but also controls transcription of genes and translation of proteins involved in microtubule dynamics. Moreover, it influences growth and proliferation of cells, as well as glucose metabolism (Pattingre et al. Through integration of intracellular signaling and growth factors, this kinase maintains the balance between protein biosynthesis and cell growth. In addition to the above mechanism, due to inactivation of the Akt kinase, the FoxO3 transcription factor is not phosphorylated. As such, it migrates to the nucleus and stimulates expression of genes coding for proteins involved in the autophagy process, i. This transcription factor is also translocated to the nucleus and stimulates expression of genes involved in the formation of autophagosomes and lysosomal biogenesis (Vodicka et al. There are two receptors in the cell membrane responsible for detection of these compounds. The Ca2+/calpain pathway the Ca2+/calpain pathway is activated under conditions of any severe changes of physiological conditions. Another factor influencing this pathway is concentration of Ca2+ ions inside the cell, which activate calpains, proteins belonging to the family of cysteine proteinases (Goll et al. They can be activated not only by Ca2+ ions transported into the cell through the calcium channel, but also by the ions liberated from endoplasmic reticulum (Gordon et al. Autophagy induction by the Ca2+/calpain pathway can be stimulated by antagonists of Ca2+ canals type L (Williams et al. They inhibit the inflow of the ions to the cell, thus, calpains are not activated. Generally, high Ca2+ levels and calpain activation negatively regulate the autophagosome formation and its fusion with lysosome (Sato-Kusubata et al. However, when the Ca2+ canals are closed, the level of Ca2+ ions is low, calpains are not activated, and the autophagy process can be initiated (Williams et al. The inositol-dependent pathway One of autophagy activation pathways is a cascade of reactions dependent on the intracellular levels of inositol. There is a link between the inositol-dependent pathway and the Ca2+/calpaine pathway. This leads to inhibition of interaction between Bcl-2 and Beclin-1, and dissociation of the phosphorylated Bcl-2 form from the Bcl-2-Beclin-1 complex. Liberated Beclin-1 can interact with hVps34, which is a prerequisite to form the Beclin-1-hVps34-hVps15 complex. The latter complex directly stimulates formation of the autophagosome (Pattingre et al.